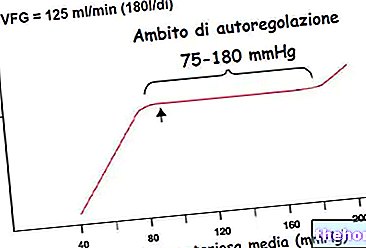
ไกลโคเจนเป็นโมเลกุลขนาดใหญ่ (มวลโมเลกุลประมาณ 400 ล้านดาลตัน) ของ α-glucose ซึ่งส่วนใหญ่มีพันธะไกลโคซิดิก α-1,4 และการแตกแขนงในอัตราส่วน 1:10 เนื่องจากพันธะไกลโคซิดิก α-1,6
ไกลโคเจนถือเป็นวัสดุสำรองและถูกย่อยสลายและสร้างใหม่อย่างต่อเนื่อง ในมวลเซลล์ทั้งหมดของร่างกาย มีไกลโคเจนประมาณ 100 กรัม ส่วนใหญ่อยู่ในตับที่เคลื่อนที่ได้ ดังนั้นจึงสามารถใช้เป็นสารสำรองสำหรับอวัยวะอื่นๆ (ไกลโคเจนในกล้ามเนื้อไม่เคลื่อนที่)
เอ็นไซม์ที่กระตุ้นการย่อยสลายและการสังเคราะห์ไกลโคเจนล้วนอยู่ในไซโตพลาสซึม ดังนั้นจำเป็นต้องมีระบบควบคุมที่ทำให้เส้นทางหนึ่งไม่ทำงานเมื่ออีกเส้นทางหนึ่งทำงาน: หากมีกลูโคสอยู่ เอ็นไซม์หลังจะถูกแปลงเป็นไกลโคเจน (แอนะโบลิซึม) ซึ่งก็คือ สำรองในทางกลับกันถ้า c "เป็นสิ่งจำเป็นสำหรับกลูโคส ไกลโคเจนจะเสื่อมโทรม (แคแทบอลิซึม)

เอนไซม์ที่เกี่ยวข้องกับการสลายไกลโคเจนเป็นส่วนใหญ่คือ ไกลโคเจนฟอสโฟรีเลส; เอนไซม์นี้สามารถแยกพันธะไกลโคซิดิก α-1,4 โดยใช้ออร์โธฟอสเฟตอนินทรีย์เป็นสารไลติก: ความแตกแยกเกิดขึ้นโดยวิธีฟอสโฟโรไลติกและได้กลูโคส 1-ฟอสเฟต
ที่ห้าหรือหกหน่วยจากจุดแตกแขนง เอ็นไซม์ไกลโคเจน ฟอสโฟรีเลส จะไม่สามารถทำงานได้อีกต่อไป ดังนั้นจึงแยกตัวออกจากไกลโคเจนและถูกแทนที่ด้วยเอ็นไซม์ deramifying ซึ่งเป็น โอน: ในตำแหน่งเร่งปฏิกิริยาของเอ็นไซม์ c "คือ" ฮิสติดีนซึ่งช่วยให้ถ่ายโอนหน่วยแซ็กคาไรด์สามหน่วยไปยังสายไกลโคซิดิกที่ใกล้ที่สุด (ฮิสติดีนโจมตีคาร์บอนตัวแรกของโมเลกุลกลูโคส) เอนไซม์ที่กล่าวถึงคือ ไกลโคซิลทรานสเฟอเรส; ในตอนท้ายของการกระทำของเอนไซม์นี้มีเพียงหน่วยกลูโคสเดียวเท่านั้นที่ยังคงอยู่ในสายโซ่ด้านข้างโดยมีคาร์บอนตัวแรกจับกับคาร์บอนที่หกของกลูโคสในสายโซ่หลักหน่วยกลูโคสสุดท้ายในสายด้านข้างจะถูกปล่อยออกมาโดยการกระทำของ "เอนไซม์ α-1,6 ไกลโคซิเดส (เอ็นไซม์นี้ถือเป็นส่วนที่สองของเอ็นไซม์ deramifying) โดยที่กิ่งก้านของไกลโคเจนอยู่ในอัตราส่วน 1:10 จากการย่อยสลายอย่างสมบูรณ์ของโมเลกุลขนาดใหญ่ เราจะได้กลูโคส 1-ฟอสเฟตประมาณ 90% และประมาณ 10% ของกลูโคส กลูโคส
การกระทำของเอ็นไซม์ดังกล่าวทำให้สามารถกำจัดสายโซ่ด้านข้างออกจากโมเลกุลไกลโคเจน การทำงานของเอ็นไซม์เหล่านี้สามารถทำซ้ำได้จนกว่าจะเกิดการเสื่อมสภาพของสายโซ่อย่างสมบูรณ์
ลองพิจารณาเซลล์ตับ กลูโคส (หลอมรวมผ่านอาหาร) เมื่อเข้าสู่เซลล์จะถูกแปลงเป็นกลูโคส 6-ฟอสเฟตและถูกกระตุ้น กลูโคส 6-ฟอสเฟต โดยการกระทำของ ฟอสโฟกลูโคมูเทสถูกเปลี่ยนเป็นกลูโคส 1 ฟอสเฟต: หลังเป็นสารตั้งต้นของการสังเคราะห์ทางชีวะที่ไม่เกิดขึ้นในทันที ในการสังเคราะห์ทางชีวสังเคราะห์จะใช้รูปแบบที่กระตุ้นของน้ำตาลซึ่งแทนด้วยน้ำตาลที่เชื่อมโยงกับไดฟอสเฟต: โดยปกติคือยูริดิลไดฟอสเฟต (UDP) กลูโคส 1- ฟอสเฟตคือ แล้วแปลงเป็น UDP-glucose เมแทบอไลต์นี้ภายใต้การกระทำของ ไกลโคเจนซินเทส ซึ่งสามารถจับ UDP-glucose กับปลายไกลโคเจนที่กำลังเติบโตโดยไม่ลดขนาด: ได้ไกลโคเจนที่ยืดยาวของหน่วยกลูโคซิดิกและได้ UDP UDP ถูกแปลงโดยเอ็นไซม์ไดฟอสโฟไคเนสที่เป็นนิวคลีโอไซด์เป็น UTP ซึ่งจะกลับสู่การไหลเวียน
การเสื่อมสภาพของไกลโคเจนเกิดขึ้นจากการกระทำของ ไกลโคเจนฟอสโฟรีเลส ซึ่งปล่อยโมเลกุลของกลูโคสและเปลี่ยนเป็นกลูโคส 1-ฟอสเฟต ต่อจากนั้น ฟอสโฟกลูโคมูเทสจะเปลี่ยนกลูโคส 1-ฟอสเฟตเป็นกลูโคส 6-ฟอสเฟต
ไกลโคเจนถูกสังเคราะห์ขึ้นเหนือสิ่งอื่นใดในตับและกล้ามเนื้อ: ในร่างกายมีไกลโคเจน 1-1.2 เฮกโตกรัมกระจายไปทั่วมวลกล้ามเนื้อ
ไกลโคเจนของไมโอไซต์เป็นตัวแทนของพลังงานสำรองสำหรับเซลล์นี้เท่านั้น ในขณะที่ไกลโคเจนที่มีอยู่ในตับก็เป็นแหล่งสำรองสำหรับเนื้อเยื่ออื่นด้วย กล่าวคือ สามารถส่งไปยังเซลล์อื่นๆ ในรูปกลูโคสได้
กลูโคส 6-ฟอสเฟตที่ได้รับในกล้ามเนื้อจากการเสื่อมสภาพของไกลโคเจนจะถูกส่งไปยังไกลโคไลซิสในกรณีที่ต้องการพลังงาน ในตับ กลูโคส 6-ฟอสเฟต จะถูกแปลงเป็นกลูโคสโดยการกระทำของ กลูโคส 6-ฟอสเฟต ฟอสฟาเตส (เอนไซม์ที่มีลักษณะเฉพาะของตับ) และถูกลำเลียงเข้าสู่กระแสเลือด
ไกลโคเจนซินเทสและไกลโคเจนฟอสโฟรีเลสทำงานบนหน่วยที่ไม่รีดิวซ์ของไกลโคเจน ดังนั้นจึงต้องมีสัญญาณของฮอร์โมนที่สั่งงานทางเดินหนึ่งและการปิดกั้นของอีกทางหนึ่ง (หรือในทางกลับกัน)
ในห้องปฏิบัติการ เป็นไปได้ที่จะยืดสายโซ่ไกลโคเจนโดยใช้ประโยชน์จากไกลโคเจน ฟอสโฟรีเลส และใช้กลูโคส 1-ฟอสเฟตในระดับความเข้มข้นสูงมาก
ในเซลล์ ไกลโคเจนฟอสโฟรีเลสเพียงเร่งปฏิกิริยาการย่อยสลายเพราะความเข้มข้นของสารเมแทบอไลต์เป็นเหมือนการเปลี่ยนสมดุลของปฏิกิริยาต่อไปนี้ไปทางขวา (กล่าวคือ ไปสู่การสลายตัวของไกลโคเจน):

เรามาดูกลไกการออกฤทธิ์ของไกลโคเจนฟอสโฟรีเลส: ออกซิเจนอะซีตัล (ซึ่งทำหน้าที่เป็นสะพานเชื่อมระหว่างหน่วยกลูโคส) จับกับไฮโดรเจนของฟอสโฟริล: ปฏิกิริยาระดับกลางเกิดจากคาร์โบเคชั่น (บนกลูโคสซึ่งก็คือทั้งหมด " แขนขา) ซึ่งฟอสโฟริล (Pi) จับตัวเร็วมาก
ไกลโคเจนฟอสโฟรีเลสต้องการโคแฟคเตอร์ซึ่งเป็นไพริดอกซอลฟอสเฟต (โมเลกุลนี้ยังเป็นปัจจัยร่วมสำหรับทรานส์อะมิเนส): มีฟอสโฟรีลโปรตอนบางส่วน (ไพริดอกซอลฟอสเฟตล้อมรอบด้วยสภาพแวดล้อมที่ไม่ชอบน้ำซึ่งพิสูจน์ว่ามีโปรตอนที่จับกับมัน) . ฟอสโฟริล (Pi) สามารถถ่ายโอนโปรตอนไปยังไกลโคเจนได้เนื่องจากฟอสโฟรีลนี้จะนำโปรตอนกลับคืนมาจากฟอสโฟรีลที่ถูกโปรตอนบางส่วนของไพริดอกซอลฟอสเฟต ความน่าจะเป็นที่ค่า pH ทางสรีรวิทยา ฟอสโฟริลจะสูญเสียโปรตอนและยังคงลดสัดส่วนลงอย่างสมบูรณ์นั้นต่ำมาก
ตอนนี้เรามาดูกันว่าฟอสโฟกลูโคมูเทสทำงานอย่างไร เอนไซม์นี้แสดงสารตกค้างของ phosphorylated serine ในบริเวณตัวเร่งปฏิกิริยา ซีรีนให้ฟอสฟอริลเป็นกลูโคส 1-ฟอสเฟต (ในตำแหน่งที่หก): กลูโคส 1,6-บิสฟอสเฟตจะเกิดขึ้นในช่วงเวลาสั้นๆ จากนั้นซีรีนจะถูกรีฟอสโฟรีเลตโดยนำฟอสโฟริลในตำแหน่งที่หนึ่ง ฟอสโฟกลูโคมิวเตสสามารถทำงานได้ทั้งสองทิศทาง เช่น การเปลี่ยนกลูโคส 1-ฟอสเฟตเป็นกลูโคส 6-ฟอสเฟต หรือในทางกลับกัน หากผลิตกลูโคส 6-ฟอสเฟต สามารถส่งโดยตรงไปยังไกลโคไลซิส ในกล้ามเนื้อ หรือเปลี่ยนเป็นกลูโคสในตับ
เอนไซม์ uridyl phosphogluco transferase (หรือ UDP กลูโคส pyrophosphorylase) เร่งปฏิกิริยาการถ่ายโอนกลูโคส 1-ฟอสเฟตไปเป็น UTP โดยยึดติดกับฟอสโฟริลเอ
เอนไซม์ที่เพิ่งอธิบายไปคือ pyrophosphorylase ชื่อนี้เกิดจากปฏิกิริยาตรงกันข้ามกับปฏิกิริยาที่เพิ่งอธิบายไปคือ pyrophosphorylation
กลูโคส UDP ที่ได้รับตามที่อธิบายไว้สามารถยืดสายโซ่ไกลโคเจนโดยใช้หน่วยโมโนแซ็กคาไรด์
เป็นไปได้ที่จะทำให้ปฏิกิริยามีวิวัฒนาการไปสู่การก่อตัวของกลูโคส UDP โดยการกำจัดผลิตภัณฑ์ที่เป็นไพโรฟอสเฟต เอนไซม์ ไพโรฟอสฟาเตส แปลงไพโรฟอสเฟตเป็นสองโมเลกุลของออร์โธฟอสเฟต (การไฮโดรไลซิสของแอนไฮไดรด์) และในการทำเช่นนั้น จะทำให้ความเข้มข้นของไพโรฟอสเฟตต่ำมากจนทำให้กระบวนการสร้างกลูโคส UDP เป็นที่ชื่นชอบทางเทอร์โมไดนามิกส์
ดังที่ได้กล่าวมาแล้ว กลูโคส UDP ด้วยการกระทำของไกลโคเจนสังเคราะห์ สามารถยืดสายโซ่ไกลโคเจนได้
การแตกแขนง (ในอัตราส่วน 1:10) เกิดจากการที่สายโซ่ไกลโคเจนประกอบด้วย 20-25 หน่วย เอ็นไซม์ที่แตกแขนง (มี "ฮิสติดีนบนไซต์ตัวเร่งปฏิกิริยา) เข้ามาแทรกแซง ซึ่งสามารถถ่ายโอนชุดของ 7 -8 ยูนิตไกลโคซิดิกต่อปลายน้ำอีก 5-6 ยูนิต: ดังนั้นจึงสร้างการแตกแขนงใหม่
ด้วยเหตุผลของต้นกำเนิดประสาทหรือหากต้องการพลังงานเนื่องจากการออกแรงทางกายภาพ อะดรีนาลีนจะถูกหลั่งออกจากต่อมหมวกไต
เซลล์เป้าหมายของ adrenaline (และ noradrenaline) คือเซลล์ตับ กล้ามเนื้อ และเนื้อเยื่อไขมัน (ในระยะหลังมีการย่อยสลายของไตรกลีเซอไรด์และการไหลเวียนของกรดไขมัน ดังนั้น กลูโคสจึงถูกผลิตขึ้นในไมโตคอนเดรีย 6 -ฟอสเฟต ส่งไปยังไกลโคไลซิสในขณะที่อยู่ใน adipocytes กลูโคส 6-ฟอสเฟตจะเปลี่ยนเป็นกลูโคสโดยการกระทำของเอนไซม์กลูโคส 6-ฟอสเฟตฟอสฟาเตสและส่งออกไปยังเนื้อเยื่อ)
มาดูกันว่าตอนนี้รูปแบบของการกระทำของอะดรีนาลีน อะดรีนาลีนจับกับตัวรับที่วางอยู่บนเยื่อหุ้มเซลล์ (ของ myocytes และ hepatocytes) และเป็นตัวกำหนดการแปลสัญญาณจากภายนอกสู่ภายในเซลล์ โปรตีน kinase ถูกกระตุ้นซึ่งทำหน้าที่พร้อมกันในระบบที่ควบคุมการสังเคราะห์และการย่อยสลายของไกลโคเจน:
การสังเคราะห์ไกลโคเจนมีอยู่สองรูปแบบ: รูปแบบ dephosphorylated (ใช้งานอยู่) และรูปแบบ phosphorylated (ไม่ใช้งาน); โปรตีน kinase phosphorylates ไกลโคเจนซินเทสและขัดขวางการทำงานของมัน
Glycogen phosphorylase สามารถมีอยู่ได้สองรูปแบบ: รูปแบบที่ใช้งานอยู่ซึ่งมี phosphorylated serine และรูปแบบที่ไม่ใช้งานซึ่ง serine ถูก dephosphorylated Glycogen phosphorylase สามารถกระตุ้นได้ด้วยเอนไซม์ ไกลโคเจน ฟอสโฟรีเลส ไคเนส. ไกลโคเจน ฟอสโฟรีเลส ไคเนสจะทำงานได้หากมีฟอสโฟรีเลตและไม่ทำงานหากขาดฟอสโฟรีเลต โปรตีนไคเนสมีไกลโคเจนฟอสโฟรีเลสไคเนสซึ่งเป็นสารตั้งต้น นั่นคือ มันสามารถกระตุ้นฟอสโฟรีเลต
เมื่อสัญญาณอะดรีนาลีนหมดลง ผลกระทบที่มีต่อเซลล์ก็ต้องจบลงด้วย: เอ็นไซม์ฟอสฟาเตสจะเข้าไปแทรกแซงโปรตีนสปีชีส์