การวิเคราะห์องค์ประกอบของโปรตีนให้ค่าเฉลี่ยดังต่อไปนี้: 55% ของคาร์บอน, 7% ของไฮโดรเจนและ 16% ของไนโตรเจน; เป็นที่ชัดเจนว่าโปรตีนต่างกัน แต่องค์ประกอบเฉลี่ยของพวกเขาแตกต่างกันเล็กน้อยจากค่าที่ระบุไว้ข้างต้น .
ตามรัฐธรรมนูญ โปรตีนเป็นโมเลกุลขนาดใหญ่ที่เกิดขึ้นจากกรด α-amino ตามธรรมชาติ กรดอะมิโนเข้าร่วมผ่านพันธะเอไมด์ซึ่งเกิดขึ้นจากปฏิกิริยาระหว่างหมู่อะมิโนของกรดเอ-อะมิโนกับคาร์บอกซิลของกรดเอ-อะมิโนอีกชนิดหนึ่ง
พันธะนี้ (-CO-NH-) เรียกอีกอย่างว่าพันธะเปปไทด์เนื่องจากจับเปปไทด์ (กรดอะมิโนรวมกัน):
หนึ่งที่ได้รับคือไดเปปไทด์เพราะประกอบด้วยกรดอะมิโนสองชนิด เนื่องจากไดเปปไทด์ประกอบด้วยหมู่อะมิโนอิสระที่ปลายด้านหนึ่ง (NH2) และคาร์บอกซิลที่ปลายอีกด้านหนึ่ง (COOH) จึงสามารถทำปฏิกิริยากับกรดอะมิโนอย่างน้อยหนึ่งตัวและขยายสายโซ่ทั้งจากด้านขวาและด้านซ้ายด้วยปฏิกิริยาเดียวกัน เห็นด้านบน
ลำดับของปฏิกิริยา (ซึ่งจริงๆ แล้ว มันไม่ง่ายขนาดนั้น) สามารถดำเนินต่อไปได้ไม่มีกำหนด: จนกว่าจะมีพอลิเมอร์ที่เรียกว่า โพลีเปปไทด์ หรือ โปรตีน. ความแตกต่างระหว่างเปปไทด์และโปรตีนเชื่อมโยงกับน้ำหนักโมเลกุล โดยปกติสำหรับน้ำหนักโมเลกุลที่สูงกว่า 10,000 เราพูดถึงโปรตีน
การรวมกรดอะมิโนเข้าด้วยกันเพื่อให้ได้โปรตีนที่มีขนาดเล็กเป็นงานที่ยาก แม้ว่าเมื่อเร็ว ๆ นี้ได้มีการพัฒนาวิธีการผลิตโปรตีนจากกรดอะมิโนแบบอัตโนมัติซึ่งให้ผลลัพธ์ที่ยอดเยี่ยม
โปรตีนที่ง่ายที่สุดจึงประกอบด้วยกรดอะมิโน 2 ตัว ตามข้อตกลงระหว่างประเทศ การเรียงลำดับกรดอะมิโนในโครงสร้างโปรตีนเริ่มจากกรดอะมิโนที่มีหมู่อะมิโนอิสระ
การเข้ารหัสสำหรับโปรตีนนี้) ซึ่งประสบปัญหาทางเคมีที่ไม่สำคัญ
เป็นไปได้ที่จะกำหนดลำดับของกรดอะมิโนผ่านการย่อยสลายของ Edman: โปรตีนทำปฏิกิริยากับฟีนิลลิโซไธโอไซยาเนต (FITC) ในขั้นต้น α-อะมิโนไนโตรเจนดับเบิ้ลโจมตีฟีนิลลิโซไธโอไซยาเนตซึ่งก่อตัวเป็นอนุพันธ์ของไทโอคาร์บามิล ต่อมาผลิตภัณฑ์ที่ได้รับจะหมุนเวียนโดยให้อนุพันธ์ฟีนิลไทโอไฮแดนโทอินซึ่งเป็นสารเรืองแสง
เอ็ดแมนได้คิดค้นเครื่องจักรที่เรียกว่าซีเควนเซอร์ ซึ่งจะปรับพารามิเตอร์โดยอัตโนมัติ (เวลา รีเอเจนต์ ค่า pH ฯลฯ) สำหรับการย่อยสลายและจัดเตรียมโครงสร้างหลักของโปรตีน (ด้วยเหตุนี้เขาจึงได้รับรางวัลโนเบล)
โครงสร้างปฐมภูมิไม่เพียงพอที่จะตีความคุณสมบัติของโมเลกุลโปรตีนได้อย่างเต็มที่ เป็นที่เชื่อกันว่าคุณสมบัติเหล่านี้ขึ้นอยู่กับโครงสร้างเชิงพื้นที่ซึ่งโมเลกุลของโปรตีนมีแนวโน้มที่จะสันนิษฐานโดยวิธีที่สำคัญ พับในลักษณะต่างๆ นั่นคือ สมมติว่าสิ่งที่ถูกกำหนดให้เป็นโครงสร้างรองของโปรตีน
โครงสร้างรองของโปรตีนกำลังกะพริบ กล่าวคือ มันมีแนวโน้มที่จะสลายตัวโดยความร้อน จากนั้นโปรตีนจะสลายตัวเองโดยสูญเสียคุณสมบัติเฉพาะหลายอย่าง นอกจากการให้ความร้อนที่สูงกว่า 70 ° C แล้ว การทำให้เสียสภาพยังสามารถเกิดจากการฉายรังสีหรือโดยการกระทำของสารตั้งต้น (เช่น จากกรดแก่)
การสลายตัวของโปรตีนอันเนื่องมาจากผลกระทบทางความร้อนนั้นสังเกตได้ เช่น โดยการให้ความร้อนแก่ไข่ขาว: จะเห็นว่ามีลักษณะเป็นวุ้นและกลายเป็นสารสีขาวที่ไม่ละลายน้ำ อย่างไรก็ตาม การเสียสภาพของโปรตีนนำไปสู่การทำลายโครงสร้างทุติยภูมิของพวกมัน แต่ปล่อยให้โครงสร้างหลักของพวกมันไม่เปลี่ยนแปลง (การรวมตัวของกรดอะมิโนต่างๆ)
โปรตีนใช้โครงสร้างระดับตติยภูมิเมื่อสายโซ่ของพวกมัน แม้ว่าจะยังคงยืดหยุ่นได้แม้ว่าจะมีการดัดโค้งของโครงสร้างทุติยภูมิ แต่ก็พับในลักษณะที่ก่อให้เกิดการจัดเรียงสามมิติที่บิดเบี้ยวในรูปทรงของร่างกายที่เป็นของแข็ง ความรับผิดชอบต่อโครงสร้างตติยภูมินั้นอยู่เหนือพันธะซัลไฟด์ทั้งหมดที่สามารถสร้างได้ระหว่างซิสเทอีน -SH ที่กระจัดกระจายไปตามโมเลกุล
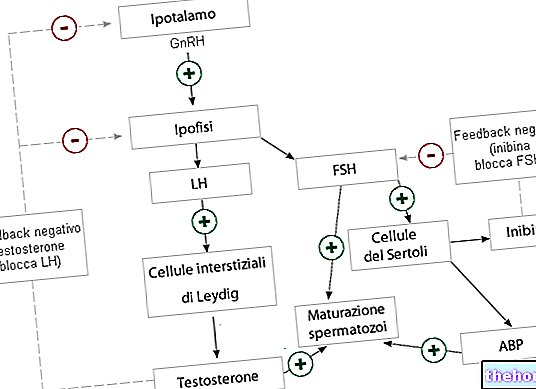
โครงสร้างควอเทอร์นารีนั้นเป็นของโปรตีนที่เกิดจากหน่วยย่อยสองหน่วยขึ้นไปเท่านั้น ตัวอย่างเช่น เฮโมโกลบินประกอบด้วยโปรตีนสองคู่ (นั่นคือในสายโปรตีนทั้งสี่) ซึ่งตั้งอยู่ที่จุดยอดของจัตุรมุขเพื่อให้เกิดโครงสร้างทรงกลม โซ่โปรตีนทั้งสี่ถูกยึดเข้าด้วยกันโดยแรงไอออนิกและ พันธะที่ไม่ใช่โควาเลนต์
อีกตัวอย่างหนึ่งของโครงสร้างควอเทอร์นารีคืออินซูลิน ซึ่งดูเหมือนจะประกอบด้วยหน่วยย่อยโปรตีนมากถึง 6 หน่วยที่จัดเรียงเป็นคู่ที่จุดยอดของรูปสามเหลี่ยมที่อยู่ตรงกลางซึ่งมีอะตอมของสังกะสีสองอะตอม
โปรตีนเส้นใย
พวกเขาเป็นโปรตีนที่มีความแข็งแกร่งบางอย่างและมีแกนยาวกว่าโปรตีนอื่น ๆ โปรตีนเส้นใยที่มีอยู่ในธรรมชาติในปริมาณที่มากขึ้นคือคอลลาเจน (หรือคอลลาเจน)
โปรตีนที่มีเส้นใยสามารถกำหนดโครงสร้างรองที่แตกต่างกัน: α-helix, β-sheet และในกรณีของคอลลาเจน, เกลียวสามชั้น; α-helix เป็นโครงสร้างที่เสถียรที่สุด รองลงมาคือ β-sheet ในขณะที่สามเกลียวที่มีความเสถียรน้อยที่สุด
ขวา ถ้าตามโครงกระดูกหลัก (เน้นจากล่างขึ้นบน) จะมีการเคลื่อนไหวคล้ายกับการขันสกรูด้านขวา ในขณะที่เกลียวเป็นของ มือซ้าย ถ้าการเคลื่อนไหวคล้ายกับการขันสกรูของสกรูมือซ้าย ในมือขวา α-เฮลิซ หมู่แทนที่ -R ของกรดอะมิโนตั้งฉากกับแกนหลักของโปรตีนและหันออกด้านนอกในขณะที่อยู่ทางซ้าย- มือ a-helices แทนที่ -R หันเข้าด้านใน a-helices ที่ถนัดขวามีความเสถียรมากกว่ามือซ้ายเพราะระหว่าง vati -R c "มีปฏิสัมพันธ์น้อยกว่าและมีอุปสรรค steric น้อยกว่า a-helices ทั้งหมดที่พบในโปรตีนคือ dextrorotal
โครงสร้างของ α-helix นั้นเสถียรโดยพันธะไฮโดรเจน (สะพานไฮโดรเจน) ที่เกิดขึ้นระหว่างหมู่คาร์บอกซิล (-C = O) ของกรดอะมิโนแต่ละชนิดและหมู่อะมิโน (-NH) พบสารตกค้างสี่ตัวในภายหลัง ลำดับเชิงเส้น
ตัวอย่างของโปรตีนที่มีโครงสร้าง α-helix คือ แฮร์เคราติน
โดยการขยายโครงสร้าง α-helix การเปลี่ยนจาก α-helix เป็น β-sheet จะดำเนินการ ความร้อนหรือความเค้นทางกลยังปล่อยให้ผ่านจาก α-helix ไปยังโครงสร้างแผ่น β
โดยปกติ ในโปรตีน โครงสร้างแผ่น β จะอยู่ใกล้กัน เนื่องจากสามารถสร้างพันธะไฮโดรเจนระหว่างสายโซ่ระหว่างส่วนต่างๆ ของโปรตีนเองได้
ในโปรตีนเส้นใย โครงสร้างโปรตีนส่วนใหญ่จัดอยู่ใน α-helix หรือ β-ชีต
โปรตีนทั่วโลก
พวกมันมีโครงสร้างเชิงพื้นที่เกือบเป็นทรงกลม (เนื่องจากการเปลี่ยนแปลงมากมายในทิศทางของสายโซ่โพลีเปปไทด์) บางส่วนของสิ่งมีชีวิตสามารถตรวจสอบย้อนกลับไปยังโครงสร้าง α-helix หรือ β-sheet และส่วนอื่น ๆ ไม่ได้มาจากรูปแบบเหล่านี้แทน: การจัดเรียงไม่ใช่แบบสุ่ม แต่มีการจัดระเบียบและทำซ้ำ
โปรตีนที่อ้างถึงตอนนี้เป็นสารที่มีโครงสร้างเป็นเนื้อเดียวกันอย่างสมบูรณ์ นั่นคือ ลำดับกรดอะมิโนบริสุทธิ์ที่รวมกัน กล่าวได้ว่าโปรตีนดังกล่าว เรียบง่าย; มีโปรตีนประกอบด้วยส่วนโปรตีนและส่วนที่ไม่ใช่โปรตีน (กลุ่มต่อมลูกหมาก) เรียกว่าโปรตีน ผัน.
ในเล็บ ในกระจกตา และในเลนส์ตา ระหว่างช่องว่างคั่นระหว่างอวัยวะบางส่วน (เช่น ตับ) เป็นต้น
โครงสร้างของมันให้ความสามารถทางกลโดยเฉพาะ มีความแข็งแรงเชิงกลสูงซึ่งสัมพันธ์กับความยืดหยุ่นสูง (เช่น ในเอ็น) หรือความแข็งแกร่งสูง (เช่น ในกระดูก) ขึ้นอยู่กับหน้าที่ที่ต้องทำ
หนึ่งในคุณสมบัติที่น่าสงสัยที่สุดของคอลลาเจนคือความเรียบง่าย: ประกอบด้วยโพรลีนประมาณ 30% และไกลซีนประมาณ 30%; กรดอะมิโนอีก 18 ชนิดต้องแบ่งโครงสร้างโปรตีน 40% ที่เหลือเท่านั้น ลำดับกรดอะมิโนของคอลลาเจนนั้นสม่ำเสมออย่างน่าทึ่ง: สำหรับทุกๆ สามสิ่งตกค้าง ลำดับที่สามคือไกลซีน
โพรลีนเป็นกรดอะมิโนแบบไซคลิกซึ่งกลุ่ม R จับกับไนโตรเจน α-อะมิโนและทำให้มีความฝืด
โครงสร้างสุดท้ายคือสายโซ่ที่ซ้ำซากซึ่งมีรูปร่างเป็น "เกลียว" ภายในสายโซ่คอลลาเจน พันธะไฮโดรเจนจะหายไป คอลลาเจนเป็น "เกลียวซ้ายมือที่มีระยะพิทช์ (ความยาวตรงกับเกลียวหนึ่งรอบ) มากกว่า" α-helix; เกลียวคอลลาเจนจะหลวมจนสายโปรตีนสามสายสามารถพันกันเป็น " เชือกเดี่ยว: โครงสร้างเกลียวสามชั้น
อย่างไรก็ตาม เกลียวสามชั้นของคอลลาเจนมีความคงตัวน้อยกว่าทั้ง α-helix และโครงสร้าง β-ชีต
มาดูกลไกการสร้างคอลลาเจนกัน พิจารณา ตัวอย่างเช่น การแตกของหลอดเลือด: การแตกนี้มาพร้อมกับสัญญาณมากมายที่มีจุดประสงค์ในการปิดหลอดเลือด ทำให้เกิดก้อน
การแข็งตัวของเลือดต้องใช้เอนไซม์พิเศษอย่างน้อยสามสิบตัว หลังจากเป็นก้อนแล้วจำเป็นต้องทำการซ่อมแซมเนื้อเยื่อต่อไป เซลล์ใกล้แผลยังผลิตคอลลาเจน ในการทำเช่นนี้ ขั้นแรกให้กระตุ้นการแสดงออกของยีน กล่าวคือ สิ่งมีชีวิตที่เริ่มต้นจากข้อมูลของยีนสามารถผลิตโปรตีนได้ (ข้อมูลทางพันธุกรรมจะถูกคัดลอกบน mRNA ซึ่งออกจากนิวเคลียสและไปถึงไรโบโซมใน ไซโตพลาสซึมซึ่งข้อมูลทางพันธุกรรมถูกแปลเป็นโปรตีน) จากนั้นคอลลาเจนจะถูกสังเคราะห์ในไรโบโซม (ดูเหมือนเกลียวซ้ายมือที่ประกอบด้วยกรดอะมิโนประมาณ 1200 ตัว และมีน้ำหนักโมเลกุลประมาณ 150,000 d) แล้วสะสมในลูเมนโดยที่ มันกลายเป็นสารตั้งต้นสำหรับเอ็นไซม์ที่สามารถดำเนินการดัดแปลงหลังการแปล (การปรับเปลี่ยนของภาษาที่แปลโดย "mRNA)" ในคอลลาเจน การดัดแปลงเหล่านี้ประกอบด้วยไฮดรอกซิเลชันของสายด้านข้างโดยเฉพาะโพรลีนและไลซีน
ความล้มเหลวของเอนไซม์ที่นำไปสู่การเปลี่ยนแปลงเหล่านี้ทำให้เกิดเลือดออกตามไรฟัน: เป็นโรคที่ทำให้หลอดเลือดแตกในขั้นต้น ฟันแตก ตามมาด้วยเลือดออกในลำไส้และเสียชีวิต อาจเกิดจากการใช้อาหารที่มีอายุยืนยาวอย่างต่อเนื่อง
ต่อจากนั้น เนื่องจากการกระทำของเอ็นไซม์อื่น การดัดแปลงอื่น ๆ เกิดขึ้นซึ่งประกอบด้วย glycosidation ของกลุ่มไฮดรอกซิลของโพรลีนและไลซีน (น้ำตาลจับกับออกซิเจนของ OH); เอ็นไซม์เหล่านี้พบได้ในบริเวณอื่นที่ไม่ใช่ลูเมน ดังนั้น โปรตีนในขณะที่ทำการดัดแปลง จะย้ายภายในเอนโดพลาสมิกเรติคูลัมไปสิ้นสุดในถุง (ถุง) ที่ปิดตัวเองและแยกออกจากเรติคูลัม: ภายในนั้นมีโปรไกลโคซิเดต -คอลลาเจนโมโนเมอร์ หลังไปถึงเครื่องมือกอลจิที่เอ็นไซม์จำเพาะรู้จักซีสเตอีนที่มีอยู่ในส่วนปลายคาร์บอกซีของโปรคอลลาเจนไกลโคซิเดต และทำให้สายโซ่ต่างๆ เข้าหากันและสร้างสะพานไดซัลไฟด์ ด้วยวิธีนี้ โซ่สามสายของ glycosidated pro-collagen ถูกเชื่อมโยงเข้าด้วยกันและนี่คือจุดเริ่มต้นที่สายโซ่ทั้งสามซึ่งแทรกซึมเข้าไปโดยธรรมชาติทำให้เกิดเกลียวสามตัว โปร-คอลลาเจนไกลโคซิเดตทั้ง 3 สายเชื่อมโยงกันไปถึง vesicle ซึ่ง สำลักตัวเอง หลุดออกจากเครื่องกอลจิ ลำเลียงโซ่ทั้งสามไปรอบ ๆ เซลล์ โดยผ่านกระแสน้ำ ไอออนกับพลาสมาเมมเบรน ไตรมิเตอร์ถูกขับออกจากเซลล์
ในพื้นที่นอกเซลล์มีเอ็นไซม์เฉพาะคือโปรคอลลาเจนเปปไทเดสซึ่งกำจัดออกจากสปีชีส์ที่ถูกขับออกจากเซลล์สามชิ้น (หนึ่งอันสำหรับแต่ละเกลียว) ของกรดอะมิโน 300 l "หนึ่งอันจากส่วนคาร์บอกซีและสาม ชิ้นส่วน (หนึ่งอันสำหรับแต่ละเกลียว) ของกรดอะมิโนประมาณ 100 ตัวจากส่วนปลายอะมิโน: ยังมีเกลียวสามตัวที่ประกอบด้วยกรดอะมิโนประมาณ 800 ตัวต่อเกลียวที่เรียกว่า โทรโปคอลลาเจน.
Tropocollagen มีลักษณะเป็นแท่งที่ค่อนข้างแข็ง ทริมเมอร์ต่างๆ เชื่อมโยงกับพันธะโควาเลนต์เพื่อให้โครงสร้างที่ใหญ่ขึ้น: ไมโครไฟเบอร์. ในไมโครไฟเบอร์ ทริมเมอร์ต่างๆ จะถูกจัดเรียงในลักษณะที่เซ microfibrils จำนวนมากเป็นการรวมกลุ่มของ tropocollagen
ในกระดูก ระหว่างเส้นใยคอลลาเจน มีช่องว่างคั่นซึ่งซัลเฟตและฟอสเฟตของแคลเซียมและแมกนีเซียมถูกสะสม เกลือเหล่านี้ยังครอบคลุมเส้นใยทั้งหมด ทำให้กระดูกแข็ง
ในเส้นเอ็น ช่องว่างคั่นระหว่างหน้าจะอุดมไปด้วยผลึกน้อยกว่าในกระดูก ในขณะที่มีโปรตีนขนาดเล็กกว่าในโทรโพคอลลาเจน ซึ่งจะทำให้เส้นเอ็นมีความยืดหยุ่น
โรคกระดูกพรุนเป็นโรคที่เกิดจากการขาดแคลเซียมและแมกนีเซียม ซึ่งทำให้ไม่สามารถตรึงเกลือในบริเวณคั่นระหว่างหน้าของเส้นใยโทรโพคอลลาเจน