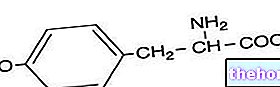
«ขั้นตอนแรกของไกลโคไลซิส
เอนไซม์ตัวแรกที่ใช้ในระยะที่สองของไกลโคไลซิสคือ กลีซาลดีไฮด์ 3-ฟอสเฟต ดีไฮโดรจีเนส; ดีไฮโดรจีเนสเป็นเอนไซม์ที่กระตุ้นการถ่ายโอนพลังงานรีดิวซ์จากโมเลกุลรีดิวซ์ที่ออกซิไดซ์ไปยังโมเลกุลอื่นที่ลดลง (ปฏิกิริยารีดอกซ์) สารตั้งต้นของเอนไซม์นี้คือ NAD (nicotidamide adenine dinucleotide) และ FAD (flavin adenin dinucleotide )
ในขั้นตอนนี้ dehydrogenase เร่งปฏิกิริยาการเปลี่ยน glyceraldehyde 3-phosphate เป็น 1,3-bisphosphoglycerate: บนไซต์ตัวเร่งปฏิกิริยาเดียวกัน กลุ่ม aldehyde จะถูกออกซิไดซ์เป็น carboxyl โดยมีผลให้ NAD + ลดลงเป็น NADH และต่อมากลุ่ม carboxyl สามารถทำได้ เพื่อสร้างพันธะแอนไฮไดรด์กับออร์โธฟอสเฟต กระบวนการแรกนั้นออกแรงมาก (ปล่อยพลังงาน) ในขณะที่ขั้นตอนที่สองนั้นออกแรงมาก (ต้องใช้พลังงาน) หากไม่มีตำแหน่งเร่งปฏิกิริยา ปฏิกิริยาทั่วโลกจะไม่เกิดขึ้น: ปฏิกิริยาแรกจะเกิดขึ้นพร้อมกับการปลดปล่อยพลังงานซึ่งจะกระจายตัวเป็นความร้อนและด้วยเหตุนี้จึงไม่สามารถนำมาใช้เพื่อสร้างพันธะแอนไฮไดรด์ได้

หลังจากการก่อตัวของ 1,3-bisphosphoglycerate เอ็นไซม์จะกลับสู่โครงสร้างเริ่มต้นและพร้อมที่จะดำเนินการกับสารตั้งต้นใหม่

แล้วก็มา ฟอสโฟกลีเซอเรต ไคเนส ซึ่งช่วยให้ถ่ายโอนฟอสฟอริลจาก 1,3-bisphosphoglycerate ไปยังโมเลกุล ADP เราได้รับ ATP (หนึ่ง ATP สำหรับแต่ละโมเลกุลของ glyceraldehyde 3-phosphate ดังนั้น ATP สองตัวสำหรับโมเลกุลกลูโคสเริ่มต้นแต่ละโมเลกุล) ซึ่งชดเชยการใช้พลังงานของ ระยะแรกของไกลโคไลซิส
แอนไอออนอาร์เซเนต (AsO43-) ส่งผลต่อวิถีไกลโคไลติกเนื่องจากสามารถแทนที่ฟอสเฟตในปฏิกิริยาแรกของระยะที่สองของไกลโคไลซิส ทำให้ 1-อาร์เซนิโอ 3-ฟอสโฟกลีเซอเรตมีความไม่เสถียรสูงทันทีที่ปราศจากตัวเร่งปฏิกิริยา ไซต์ไฮโดรไลซ์ปล่อย "สารหนูกลับสู่การไหลเวียน ดังนั้นสารหนูจึงเลียนแบบการกระทำของฟอสเฟตและเข้าสู่ไซต์ตัวเร่งปฏิกิริยา: ในที่ที่มีสารหนูปฏิกิริยาที่สร้าง ATP (จาก 1,3-bisphosphoglycerate ถึง 3-phosphoglycerate) จึงไม่เกิดขึ้นเนื่องจาก 3-phosphate glyceraldehyde ถูกแปลงเป็น 3-phosphoglycerate โดยตรง หากไม่มี ATP เซลล์ก็จะตาย (พิษของกรดอาร์เซนิก)
ในปฏิกิริยาที่สามของเฟสออกซิเดชัน 3-phosphoglycerate จะถูกแปลงเป็น 2-phosphoglycerate โดยการกระทำของ ฟอสโฟกลีเซอเรตมิวเตส; ปฏิกิริยานี้เกี่ยวข้องกับตัวกลาง 2,3-bisphosphoglycerate

ในขั้นตอนต่อไป เอนไซม์จะเข้าไปแทรกแซง อีโนเลส ซึ่งสามารถกระตุ้นการกำจัดโมเลกุลของน้ำออกจากโครงกระดูกคาร์บอนของ 2-phosphoglycerate เพื่อให้ได้ pyrivated phosphoenol (PEP)

PEP มีศักยภาพในการถ่ายโอนฟอสฟอริลสูง: ถ่ายโอนผ่านการกระทำของเอนไซม์ ไพรูเวท ไคเนสฟอสฟอริลกับ ADP เพื่อให้ ATP ในขั้นตอนที่ห้าของระยะที่สอง รับไพรูเวต

2-phosphoglycerate และ 3-phosphoglycerate มีกำลังการถ่ายโอนต่ำของ phosphoryl ดังนั้น เพื่อให้ได้ ATP จากโมเลกุลเหล่านี้ 3-phosphoglycerate จะถูกแปลงเป็น 2-phosphoglycerate ระหว่าง glycolysis เนื่องจากได้มาจาก PEP ซึ่งเป็น ชนิดที่มีศักยภาพในการถ่ายโอนสูง
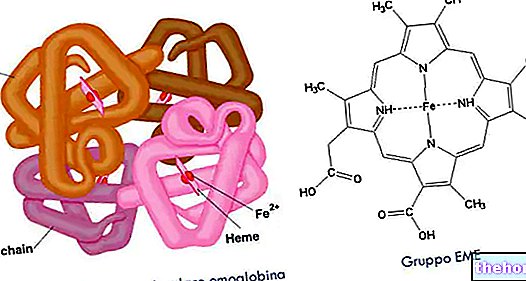
ก่อนดำเนินการต่อ ให้เปิดวงเล็บที่ 2,3-bisphosphoglycerate หลังมีอยู่ในทุกเซลล์ที่ glycolysis เกิดขึ้นในความเข้มข้นต่ำมาก (เป็นตัวกลางของปฏิกิริยาที่สามของระยะที่สองของ glycolysis) ในเม็ดเลือดแดง ในทางกลับกัน 2,3-bisphosphoglycerate มีความเข้มข้นคงที่ที่ 4-5 mM (ความเข้มข้นสูงสุด) เนื่องจากพวกมันมีสมบัติทางเอนไซม์ซึ่งมีหน้าที่ในการผลิต ในเม็ดเลือดแดง มีการเบี่ยงเบนจาก glycolysis เพื่อผลิต 2,3-bisphosphoglycerate: 1,3-bisphosphoglycerate จะถูกแปลงเป็น 2,3-bisphosphoglycerate โดยการกระทำของ bisphosphoglycerate มิวเทส (เม็ดเลือดแดง) และ 2,3-bisphosphoglycerate โดยการกระทำของ bisphosphoglycerate ฟอสฟาเตส (เม็ดเลือดแดง) กลายเป็น 3-phosphoglycerate จากนั้นในเม็ดเลือดแดง ส่วนหนึ่งของ 1,3-bisphosphoglycerate ที่ได้จาก glycolysis จะถูกแปลงเป็น 2,3-bisphosphoglycerate ซึ่งจะกลับไปสู่เส้นทางไกลโคไลติกเป็น 3-phosphoglycerate ในการทำเช่นนั้น ขั้นตอนที่สามของระยะออกซิเดชันของ glycolysis ที่ ATP ได้มา ปริมาณของ ATP ที่สูญเสียไปคือราคาที่เม็ดเลือดแดงยินดีจ่ายเพื่อรักษาความเข้มข้น 2,3-bisphosphoglycerate ที่เซลล์เหล่านี้ต้องการเพราะส่งผลต่อความสามารถของ "เฮโมโกลบินในการจับ" ออกซิเจน
เราได้เห็นแล้วว่าในปฏิกิริยาแรกของระยะที่สองของไกลโคไลซิส NAD + จะลดลงเป็น NADH แต่มีความจำเป็นที่หลังจากได้ไพรูเวตแล้ว NADH จะถูกแปลงกลับเป็น NAD +: สิ่งนี้เกิดขึ้นได้ด้วยการหมักแลกติก (ได้แลคเตท) หรือโดยการหมักด้วยแอลกอฮอล์ (pyruvate decarboxylase ซึ่ง decarboxylates pyruvate และ dehydrogenase ซึ่งก่อให้เกิดเอทานอล) การหมักไม่เกี่ยวข้องกับออกซิเจน (anaerobes)
เนื่องจากการหมักแลคติก กรดแลคติกหากกำจัดไม่เพียงพอจะสะสมในกล้ามเนื้อและปล่อย H + ทำให้เกิดการหดตัวของกล้ามเนื้อโดยไม่สมัครใจและดังนั้นจึงเป็นตะคริว กล้ามเนื้อที่มีความเครียดสูงสามารถมีค่า pH ขั้นต่ำ 6.8 .
ผ่านวงจร Cori ส่วนหนึ่งของความเหนื่อยล้าของกล้ามเนื้อจะถูกส่งไปยังตับเมื่อกล้ามเนื้อมีมากเกินไป สมมติว่ากล้ามเนื้อทำงานโดยไม่มีการจ่ายออกซิเจน (ผิดสมมติฐาน): หากกล้ามเนื้อทำงานในระดับปานกลาง ATP ที่จำเป็นสำหรับการหดตัวนั้นมาจาก glycolysis เท่านั้น หากกิจกรรมของกล้ามเนื้อเพิ่มขึ้นและจำเป็นต้องมี ATP เพิ่มเติม ให้เร่งการเผาผลาญแอโรบิก แลคเตทซึ่งถูกกำจัดออกไปเป็นกลูโคส ในความเป็นจริง กล้ามเนื้อใช้ประโยชน์จากเมแทบอลิซึมของแอโรบิก: หากมีออกซิเจนเพียงพอ กล้ามเนื้อจะทำงานได้ เหนือสิ่งอื่นใด ATP ที่มาจากเมตาบอลิซึมแบบแอโรบิก และเมื่อไม่มีออกซิเจนแล้ว เมแทบอลิซึมแบบไม่ใช้ออกซิเจนจะถูกเร่งผ่านวงจร Cori วัฏจักรนี้อนุมานว่าแลคเตทถูกถ่ายโอนจากกล้ามเนื้อไปยังตับโดยการใช้พลังงานจะทำให้เกิดกลูโคสมากขึ้นซึ่งจะกลับไปสู่กล้ามเนื้อ ผ่านวงจรนี้ ส่วนหนึ่งของ ATP ที่บริโภคในกล้ามเนื้อจะได้รับจากตับซึ่ง ผ่านกระบวนการสร้างกลูโคนีเจเนซิส สามารถผลิตกลูโคสที่กล้ามเนื้อสามารถใช้เพื่อรับเอทีพีได้
เมแทบอลิซึมของกลูโคสที่อธิบายจนถึงตอนนี้ไม่รวมออกซิเจน แต่เมแทบอลิซึมของกลูโคสแบบแอโรบิกช่วยให้ได้รับ ATP ในปริมาณที่สูงกว่าที่ได้รับจากวิถีไกลโคไลติก 17-18 เท่า ดังนั้นเมื่อเซลล์สามารถเลือกระหว่างแอโรบิกและแอโรบิกได้ anaerobe โปรดปรานอดีต
ในการเผาผลาญแบบแอโรบิก ไพรูเวตเข้าสู่ไมโตคอนเดรียซึ่งผ่านการเปลี่ยนแปลงและในที่สุดก็ได้รับคาร์บอนไดออกไซด์และน้ำ ด้วยวิธีนี้จะได้เอทีพี 34 โมเลกุลสำหรับแต่ละโมเลกุลของกลูโคสที่เสื่อมโทรม