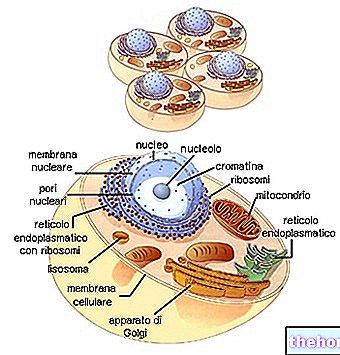
ความสำคัญของไมโอซิส
ในบริบทของสิ่งมีชีวิตหลายเซลล์ จำเป็นที่ทุกเซลล์ (เพื่อไม่ให้รับรู้ซึ่งกันและกันว่าเป็นสิ่งแปลกปลอม) มีมรดกทางพันธุกรรมที่เหมือนกัน ซึ่งทำได้โดย mitosis แบ่งโครโมโซมระหว่างเซลล์ลูกสาว ซึ่ง "ความเท่าเทียมกันของพันธุกรรม" ข้อมูลได้รับการประกันโดยกลไกการจำลองซ้ำของ DNA ในความต่อเนื่องของเซลล์ที่เปลี่ยนจากไซโกตไปยังเซลล์สุดท้ายของสิ่งมีชีวิต ในสิ่งที่เรียกว่าโซมาติกไลน์ของการสร้างเซลล์
อย่างไรก็ตาม หากใช้กลไกเดียวกันในรุ่นลูกหลานทั้งสายพันธุ์ก็มีแนวโน้มที่จะประกอบด้วยบุคคลที่มีความเท่าเทียมกันทางพันธุกรรม การขาดความแปรปรวนทางพันธุกรรมดังกล่าวอาจทำให้ความอยู่รอดของสายพันธุ์ลดลงได้ง่ายเมื่อสภาพแวดล้อมเปลี่ยนแปลง ดังนั้น จึงมีความจำเป็น ว่าสปีชีส์ ในบริบทของความแปรปรวนของสารพันธุกรรมที่ยอมรับ อาจก่อให้เกิดการแบ่งประเภทใหม่ การผสม ไม่ใช่ในบริบทของสิ่งมีชีวิตเดียว แต่ในการผ่านจากรุ่นหนึ่งไปยังอีกรุ่นหนึ่ง สิ่งนี้ทำได้โดยปรากฏการณ์ทางเพศและกลไกการแบ่งเซลล์โดยเฉพาะที่เรียกว่าไมโอซิส

ไมโอซิสคืออะไร
ไมโอซิสเกิดขึ้นเฉพาะในเซลล์สืบพันธุ์ เมื่อการแบ่งไมโทติคแบบยาวๆ ได้เพิ่มจำนวนเซลล์สืบพันธุ์ที่มีอยู่อย่างเพียงพอ เซลล์สืบพันธุ์จะเข้าสู่ไมโอซิส จึงเป็นการเตรียมเซลล์สืบพันธุ์ gametes ผสานเข้ากับการปฏิสนธิรวมวัสดุโครโมโซมของพวกมัน ถ้าเซลล์สืบพันธุ์เป็นแบบดิพลอยด์ เช่นเดียวกับเซลล์อื่นๆ ของสิ่งมีชีวิต การรวมตัวของพวกมันในไซโกตจะทำให้เด็กมีมรดก 4n เซลล์ สิ่งเหล่านี้จะให้ลูก 8 คนเป็นต้น
เพื่อรักษาจำนวนโครโมโซมของสปีชีส์ให้คงที่ gametes จะต้องเป็นเดี่ยวนั่นคือด้วยหมายเลข n แทนที่จะเป็น 2n ของโครโมโซม สิ่งนี้ทำได้ด้วยไมโอซิส
ไมโอซิสสามารถเข้าใจได้ว่าเป็นการสืบทอดของไมโทติคสองดิวิชั่นโดยไม่ทำให้เกิดการซ้ำซ้อน
ในแต่ละดิวิชั่นที่ต่อเนื่องกันสองดิวิชั่น ซึ่งสร้างเซลล์เดี่ยวสี่เซลล์จากเซลล์สืบพันธุ์แบบดิพลอยด์ มีการสืบทอดของโพรเฟส เมตาเฟส แอนาเฟส เทโลเฟส และไซโตไดเรซิส
อย่างไรก็ตาม คำทำนายของการแบ่งเซลล์แบบมีโอติกแรกนั้นซับซ้อนเป็นพิเศษ ทำให้เกิดช่วงเวลาต่อเนื่องกันซึ่งใช้ชื่อตามลำดับของเลปโตทีน ไซโกทีน ปาคีทีน ไดโพทีน และไดอะคิเนซิสตามลำดับ
เราพิจารณาโมเมนต์เหล่านี้ทีละคู่ตามพฤติกรรมของโครโมโซมคู่เดียว
เลปโตทีน. เป็นจุดเริ่มต้นของไมโอซิส โครโมโซมเริ่มมองเห็นกันยังไม่เป็นเกลียวมาก
ไซโกทีน มีการจำแนกโครโมโซมได้ชัดเจนยิ่งขึ้น และมองเห็นโครโมโซมที่คล้ายคลึงกันเข้ามาใกล้ยิ่งขึ้น (โปรดจำไว้ว่าเส้นใยที่มีแนวโน้มเข้าหาขนานกันคือ 4 โครมาทิดสำหรับโครโมโซมที่คล้ายคลึงกันแต่ละอัน)
ปาคีทีน. ฟิลาเมนต์สีโครมาทิดิกสี่เส้นยึดติดตลอดความยาว แลกเปลี่ยนจังหวะ โดยการแตกและเชื่อม
ไดโพลทีน เมื่อเกลียวและความหนาเพิ่มขึ้น โครโมโซมมีแนวโน้มที่จะแยกความแตกต่างออกจากกัน: โดยเซนโทรเมียร์แต่ละอันจะเชื่อมเข้าด้วยกันเป็นเกลียวคู่
จุดที่เกิดการแลกเปลี่ยนโดยการทำลายและการเชื่อม (chiasma) ยังคงยึดเส้นใย (chromonemes) ไว้ด้วยกันในส่วนต่างๆ โครโมนทั้งสี่ที่เชื่อมต่อเป็นคู่โดยเซนโทรเมียร์และยึดเกาะกันอย่างหลากหลายในไคอัส ก่อตัวเป็นเทโทรด
ไดอะซิเนซิส tetrads มีแนวโน้มที่จะจัดเรียงตัวเองที่เส้นศูนย์สูตรของแกนหมุน เยื่อหุ้มนิวเคลียสหายไป การแยก centromeres เริ่มต้นขึ้น เมื่อเป็นเช่นนี้โครโมโซมซึ่งรวมกันอยู่ใน chiasm แล้วแยกออกจากกัน
หลังจากเมตาเฟสถัดไป เซนโตรเมียร์ทั้งสอง (ยังไม่เพิ่มเป็นสองเท่า) จะย้ายไปยังขั้วตรงข้ามของแกนหมุน
ตามด้วยแอนาเฟส เทโลเฟส และไซโตไดเรซิสของดิวิชันที่ 1 ตามลำดับอย่างรวดเร็ว และหลังจากนั้นในดิวิชันที่สองในทันที
ในขณะที่หลังจากเมตาเฟสของดิวิชั่นแรก เซนโทรเมียร์จะย้ายไปยังขั้วของแกนหมุนที่ลากเส้นใยสองเส้น ในเมตาเฟสที่สอง แต่ละเซนโตรเมียร์จะเพิ่มเป็นสองเท่า เซลล์ทั้งสองที่เกิดจากการแบ่งส่วนแรกได้รับ n centromeres ที่มี 2n filaments แต่การแบ่งตัวที่ตามมาทำให้ได้ 4 เซลล์ แต่ละเซลล์มี n filaments (กล่าวคือ ณ จุดนี้ n โครโมโซม)
แบบแผนทั่วไปนี้อธิบายปรากฏการณ์ที่แตกต่างและขนานกันสามประการ:
- การลดลงของชุดโครโมโซมจากดิพลอยด์ (2n) ของ "สิ่งมีชีวิตเป็น" เดี่ยว (n) ของ gamete
- การแสดงที่มาแบบสุ่มของเซลล์สืบพันธุ์ของโครโมโซมหนึ่งหรืออีกโครโมโซมที่มีต้นกำเนิดจากมารดาหรือบิดา
- การแลกเปลี่ยนสารพันธุกรรมระหว่างโครโมโซมที่คล้ายคลึงกันของแหล่งกำเนิดของบิดาและมารดา (ด้วยการผสมสารพันธุกรรมไม่เพียง แต่ในระดับของโครโมโซมทั้งหมด แต่ยังอยู่ภายในโครโมโซมด้วย)